
Descrizione e diagnosi di un processo ecopatologico planetario
di Warren M. Hern – University of Colorado
L’uomo è un’apparizione
Eliot Porter, April 5, 1989 Tesuque, New Mexico
ESPOSIZIONE DEL PROBLEMA
La caratteristica più sorprendente della popolazione umana nel ventesimo secolo è la sua crescita rapida e senza freni. Eppure questo fenomeno non è nuovo. Si sono verificati diversi scatti nella crescita della popolazione umana. Uno cominciò con la fine del Paleolitico e l’inizio della rivoluzione agricola; un altro avvenne all’inizio della rivoluzione industriale (Deevey, 1960; Hassan, 1981). A partire dal 1990, negli ultimi quarant’anni si sono aggiunti più uomini alla popolazione mondiale totale rispetto ai precedenti tre milioni di anni (Keyfitz, 1989). Tra il 7 e l’8% di tutti gli esseri umani mai nati sono vivi oggi (Weeks, 1989). Fino a poco tempo fa, il tasso di crescita della popolazione umana era in aumento, il che significa che superava addirittura la crescita esponenziale (Demeny, 1986; Bartlett, 1978). Mentre l’attuale tasso di crescita ha rallentato leggermente, rimane esponenziale a circa l’1,7% all’anno con un tempo di raddoppio della popolazione di circa 40 anni. Vi è un ampio disaccordo sulle prospettive di crescita futura della popolazione umana e se, quando e in quali circostanze smetterà di crescere.
Ad accompagnare questo rapido incremento della popolazione è stato il ritmo crescente della distruzione umana dell’ecosistema globale. Gli assalti umani su piccola scala all’ambiente hanno avuto un impatto minimo o nullo durante il primo Pleistocene, sebbene gli impatti locali e regionali iniziarono a vedersi nel tardo Pleistocene e nel Neolitico (Martin, 1980; Angel, 1975; Darby, 1956). A partire dall’età industriale, vaste porzioni di ecosistemi precedentemente intatti che avevano prima sperimentato poche o nessuna perturbazione umana sono state distrutte. Mentre il 20° secolo volge al termine, le attività umane stanno seriamente e forse irreversibilmente distruggendo le caratteristiche dell’ecosistema globale che sono necessarie per la conservazione delle specie umane e delle altre forme di vita. I livelli globali di anidride carbonica atmosferica, per esempio, sono aumentati del 25% nel corso dell’ultimo secolo (Mooney, 1988). Tre degli anni più caldi degli ultimi 100 anni si sono verificati negli anni ’80 (Schneider, 1989). Severe riduzioni stagionali degli strati di ozono sull’Antartide, e forse persino sull’Artico, sono questioni di grande interesse per gli scienziati dell’atmosfera (Graedel & Crutzen, 1989). Lo smog fotochimico è diventato un problema permanente nella maggior parte delle principali città del mondo e le “megalopoli” stanno iniziando a contenere la maggior parte degli abitanti della terra (Dogon e Kasarda, 1988). Ogni anno vengono bruciate aree enormi di foreste pluviali del mondo e questi incendi producono strati di fumo che coprono decine di migliaia di chilometri quadrati. Innumerevoli specie si sono estinte come risultato diretto dell’attività umana, e il tasso di estinzioni è in aumento (Ziswiler, 1967; Hoage, 1985; Wilson, 1988). Wilson (1990) stima che, al tasso attuale di distruzione dell’ecosistema, ben il 25% di tutte le specie viventi si estinguerà entro i prossimi cinquant’anni. Oggi un capitano di una petroliera può spazzare via un intero ecosistema; incidenti di questo tipo riempiono i titoli quotidiani. Allo stesso tempo, c’è una crescente consapevolezza del nostro ecosistema globale e della necessità di preservarlo (SCEP, 1970, Boulding, 1973, Ehrlich et al, 1977; Lovelock, 1979; Russell, 1983; Myers, 1984; Rambler et al, 1989: Sahtouris, 1989; Daly & Cobb, 1989).
La popolazione umana è stata spesso paragonata a una sorta di malattia planetaria (Gregg, 1955, Eisley, 1961, Russell, 1983; Odum, 1989). Il geologo Peter Flawn, parlando agli studenti della Northwestern University nel 1970, disse che la crosta terrestre ha una malattia della pelle, causata da microbi che infettano la sua crosta e che la malattia è un uomo (Flawn, 1970). Le viste schematiche degli insediamenti umani hanno una certa somiglianza con alcuni tipi di lesioni patologiche, inclusi vari tipi di dermatiti. Flawn e altri osservatori stimolano la ricerca di altri modelli patologici. Il libro di Lewis Mumford (1961), La città nella storia, è disseminato di riferimenti a grandi città come processi patologici. Geddes (1915), includeva uno schema di città che includeva “patholopolis”. Poiché la crescita e l’invasività spettacolari sono caratteristiche eccezionali della popolazione umana, la somiglianza della specie umana con un processo cancerogeno viene subito in mente, specialmente da un medico. Una visione schematica della crescita di Londra dal 1800 al 1955 non assomiglia ad altro che a un tumore in espansione, invasivo, metastatico e maligno (Johnson, 1972, Figura 1).
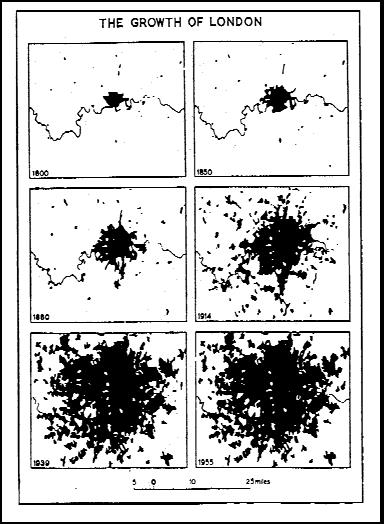
FIGURA 1. La crescita di Londra, 1800-1955 (fonte: Johnson, 1972).
Un modello quasi identico è visto nella crescita di Baltimora dal 1800 al 1950 (Gist & Fava, 1964, Figura 2, Hoover, 1948).

FIGURA 2. La crescita di Baltimora, 1800-1950 (fonte: Gist & Fava, 1964).
Caratteristiche delle popolazioni e delle comunità umane
Gli esseri umani possono essere studiati nello stesso modo in cui studiamo altre specie e organismi. Entro certi limiti, gli umani sembrano condividere le stesse caratteristiche ed essere soggetti alle stesse leggi fisiche e biologiche delle altre specie. Le curve di sopravvivenza degli umani, per esempio, possono essere paragonate a specie diverse come il cervo dalla coda nera (Taber & Dasman, 1957), i moscerini della frutta e le ostriche (Deevey, 1960). Le piramidi di popolazione possono riflettere le distribuzioni relative all’età di popolazioni umane o di altre popolazioni (Odum, 1971). Esistono, infatti, diversi tipi di distribuzione delle età tra le società umane. Una delle più comuni scoperte in tutto il mondo in questo momento è quella di una popolazione giovane e in rapido sviluppo con fino al 50% della popolazione in età riproduttiva (Hern, 1977; 1988). Le curve di crescita nelle popolazioni di varie specie tendono a livellarsi (Odum, 1971), ma alcune fasi della crescita della popolazione possono essere estremamente rapide. La specie umana ha mostrato quest’ultimo modello sin dalla sua origine con un tasso di crescita marcatamente aumentato negli ultimi secoli (Hassan, 1981, Figura 3).
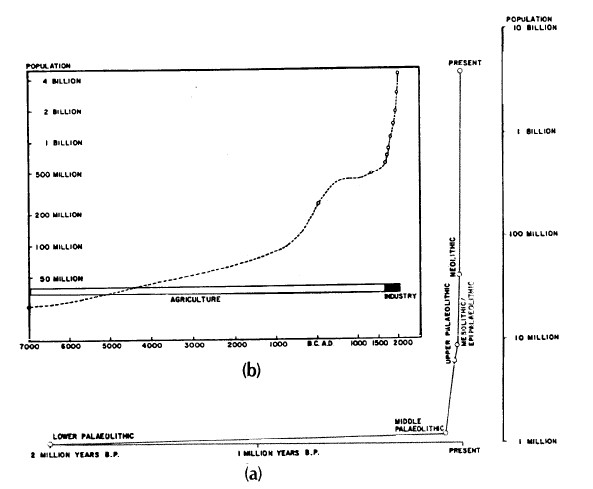
FIGURA 3. a) La crescita della popolazione umana negli ultimi due mil. di anni. (Hassan, 80) b) Modello di crescita della popolazione negli ultimi 9.000 anni. Si vede chiaramente la accelerazione del tasso di crescita della popolazione e il drammatico aumento della dimensione della popolazione mondiale negli ultimi secoli (Hassan, 78; Thomlinson 65)
Studi di biologia della popolazione mostrano ripetutamente che le specie i cui esemplari crescono rapidamente sono soggette a forti fluttuazioni delle dimensioni della popolazione e della vitalità, come ad esempio nel cervo del Kaibab Plateau (Rasmussen, 1941, Leopold, 1943; Figura 4), nel tripide adulto (Davidson & Andrewartha, 1948), e nella cardiaspina albitextura che si nutre di alberi di eucalipto (Clark, 1964). Gli incidenti demografici capitano sovente in popolazioni in rapida crescita che eccedono le capacità di carico dei loro ecosistemi o delle forniture alimentari locali. Margalef (1968) illustra questo principio con il suo diagramma di un circuito di feedback negativo (Figura 5).
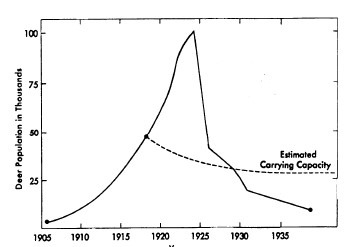
FIGURA 4. Crescita e calo della popolazione del cervo del Kaibab Plateau (Fonte: Boughey, 1973)
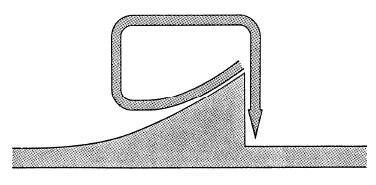
FIGURA 5. Ciclo di feedback negativo. (Fonte: Margalef, 1968)
Alcuni crolli della popolazione potrebbero non comportare alcun recupero della popolazione originaria, oppure oscillare attorno a una media che sembra essere determinata dalla capacità di carico di un dato ecosistema (Boughey, 1973, Figura 6).
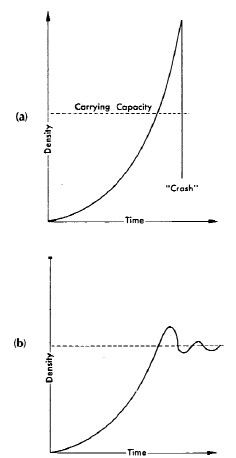
FIGURA 6. Aumento della popolazione, overshoot, crash o ritorno al livello di capacità di carico con oscillazioni smorzate (Fonte: Boughey, 1973). a) Un esempio teorico di aumento della popolazione che ha superato la capacità di carico, con conseguente incidente. b) Un esempio teorico di aumento della popolazione con una curva di forma J che ha leggermente superato il limite e oscilla attorno al livello della capacità di carico.
Le relazioni predatore-preda possono produrre fluttuazioni della dimensione della popolazione in una serie di oscillazioni collegate, come nel caso classico delle popolazioni di razze di lepri e di lince (Figura 7; MacLulich, 1937). Una singola relazione predatore-preda porta a brusche oscillazioni nelle dimensioni della popolazione, ma relazioni di predatore-preda più complesse portano a oscillazioni smorzate che tendono ad essere uniformi su tempi medi (Figura 8, Wilson & Bossert, 1971).
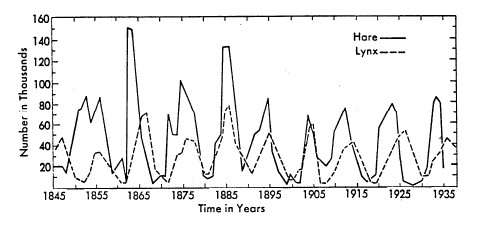
FIGURA 7. Rapporto predatore-predatore di lince e lepri con le ciaspole, che mostrano oscillazioni collegate dei livelli di popolazione (Fonte: Boughey, 1973).
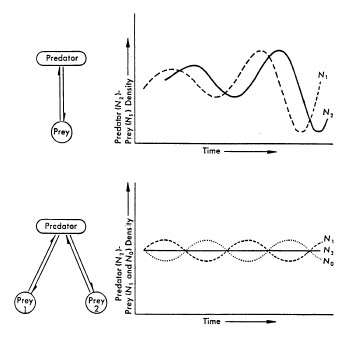
FIGURA 8. Modello di relazioni preda-predatore singolo e multiplo (Fonte: Boughey, 1973).
Un concetto importante nella biologia della popolazione è che le popolazioni tendono a mantenersi in una serie di oscillazioni smorzate. La digressione da questo modello si traduce in una grave instabilità e persino nell’estinzione.
Parte dell’instabilità e vulnerabilità all’estinzione sembra essere correlata a stress che si sviluppano in condizioni di elevata densità di popolazione e di grave affollamento, come dimostrato dai ben noti esperimenti di John Calhoun (1962) e John Christian (1980). L’affollamento conduce a patologie sociali e biologiche che si traducono in alta mortalità e ridotta sopravvivenza del gruppo (Cassel, 1971; Aaby et al, 1983).
Una caratteristica importante della popolazione umana storica è stata la capacità di aggirare questi stress attraverso la suddivisione in gruppi, l’emigrazione e la colonizzazione di nuovi territori non sfruttati. Di conseguenza, migliaia di anni di migrazione intercontinentale e intracontinentale hanno provocato la colonizzazione permanente di praticamente ogni parte dell’ecosistema terrestre globale da parte degli esseri umani e lo sviluppo di insediamenti antropici colossali e in rapida crescita che avvolgono e inghiottono gli ecosistemi adiacenti. Le colonie circostanti, o sobborghi, diventano parti indistinguibili della distesa urbana.
Mentre le strategie di migrazione e colonizzazione erano disponibili negli ultimi 10.000 anni di storia umana, divennero indisponibili quando la rapida crescita della popolazione iniziò a verificarsi. La migrazione e la concentrazione della popolazione sono in direzione delle grandi città dove lo stress di alta densità sviluppa nuove urgenze e intensità per gli individui.
Le popolazioni umane hanno risposto alle pressioni della sovrappopolazione con una varietà di strategie diverse dalla migrazione, compresa la predazione e l’intensificazione agricola (Abernethy, 1979; Sahlins, 1961; Cohen, 1977).
Comunità umane
Una caratteristica sorprendente delle comunità umane è che stanno diventando indistinguibili nell’aspetto. Le antiche comunità umane spesso esibivano modelli unici di architettura e residenza che erano caratteristici e specifici di una particolare cultura o area (Jordan & Rowntree, 1986). I confini delle città erano accuratamente delimitati dall’avere mura cittadine. Le città venivano a volte collocate per ragioni difensive in siti in cui la topografia fisica richiedeva questa pratica o forniva i mezzi con cui veniva realizzata, come nel caso delle città collinari italiane (Carver, 1979). In tempi più recenti, i confini delle comunità di tutto il mondo sono diventati indistinti mentre stanno diventando più estesi.
I pianificatori urbani e gli studenti delle comunità umane hanno fatto riferimento per molti anni ai loro soggetti in termini aggregati. La raffigurazione di Gruen (1973) dell'”organismo urbano” è mostrata nella Figura 9. Lewis Mumford (1961) scrisse che le antiche città potevano essere datate nella loro crescita quasi come gli anelli degli alberi dalla costante espansione del loro muro esterno. Ora, con il fenomeno della “conurbazione”, come fu chiamato da Patrick Geddes (1915), il nuovo «… tessuto urbano era meno differenziato del vecchio, presentava una vita istituzionale povera, mostrava meno segni di nucleazione sociale e tendeva ad aumentare di dimensioni, blocco per blocco, strada per strada, ‘sviluppo’ per ‘sviluppo’, senza alcuna individualità di forma e, cosa più notevole di tutte, senza limiti quantitativi» (Mumford, 1961).
Nel discutere i cambiamenti dell’ecosistema, compresi i cambiamenti atmosferici e vegetazionali, portati dai grandi insediamenti urbani, Wolman (1965) parla del “metabolismo della città”. Il fenomeno dell'”isola di calore urbana” è ben noto e può contribuire in modo significativo al processo di riscaldamento globale (Jordan & Rowntree, 1986).
Le città hanno dappertutto esibito un processo di “de-differenziazione” a livello di comunità, diventando superorganismi senza forma, invadendo e distruggendo l’ambiente naturale circostante e espandendosi a un ritmo rapido. La Figura 10, dei Centri per l’ambiente urbano di Gruen (1973), offre un’illustrazione fantasmagorica di questa idea. Secondo McHarg (1969), la città moderna è un deserto ecologico, ostile alle forme umane e ad altre forme di vita, dove nulla cresce.

FIGURA 10. “La città divora la campagna”. (Fonte: Gruen, 1973).
Modelli di processi patologici
Se percepiamo la specie umana come qualcosa che è distruttivo per gli altri e persino per gli ecosistemi in cui viviamo, e se iniziamo a cercare analogie o confronti con altri organismi o processi patologici per aiutarci a capire il processo a cui stiamo assistendo e di cui stiamo causando il verificarsi, dobbiamo trovare un’ipotesi che spieghi la realtà e preveda eventi. Questo è lo scopo di un’ipotesi. La nostra ipotesi deve essere quella che, entro limiti ragionevoli, ci consenta di spiegare una serie di eventi e osservazioni che finora sembravano non correlati o collegati in un modo che non è ancora chiaro per noi. In questo caso, deve essere un’ipotesi che include aspetti del comportamento umano e della biologia, nonché cambiamenti antropogenici nell’ecosistema (Cassel, 1964). Dobbiamo essere pronti a esplorare ogni possibilità.
I modelli matematici lineari hanno importanti limiti nella descrizione di sistemi di grandi dimensioni per i quali abbiamo informazioni parziali o inaffidabili e sono particolarmente carenti nel descrivere i sistemi biologici. Ad esempio, il General Circulation Model (GCM) (Schneider, 1989) è un modello matematico altamente sofisticato che descrive i cambiamenti climatici globali derivanti dall’accumulo di gas serra, ma tali modelli non spiegano l’ampia varietà di fenomeni biologici o di altri fenomeni geomorfologici che accompagnano o derivano da attività umane (Sagan, et al, 1981). Il GCM può essere esplicativo per alcuni fenomeni come i cambiamenti climatologici, ma non per la maggior parte dei cambiamenti globali, e non ci aiuta con l’inferenza causale a determinare perché il cambiamento ecologico globale sta accadendo tranne che in un senso molto limitato.
La difficoltà di usare i modelli per confrontare gli esseri umani con altre specie o processi è che gli esseri umani hanno una cultura, che consente la trasmissione della conoscenza da una generazione all’altra, a volte per migliaia di anni e migliaia di generazioni. La cultura umana ci dà la capacità di adattarci alle circostanze e agli ambienti inospitali in modi non disponibili per altre specie. Non siamo, infatti, soggetti ad alcuni degli stessi limiti e vincoli biologici che limitano le azioni e la crescita di altre specie.
Il confronto tra la specie umana e un processo patologico è pieno di rischi. Se facciamo il paragone della popolazione umana con un processo patologico come fece il professor Flawn, corriamo il rischio che non sia appropriato a causa delle differenze di scala e delle differenze di processo. Come si può confrontare la specie umana con la dermatite? La dermatite può essere causata da una varietà di fattori inclusi microbi (batteri o funghi), processi autoimmuni o allergici, sostanze tossiche o altri agenti fisici o da disturbi psicogeni. La dermatite non diventa spesso invasiva e di solito non minaccia il suo ospite. Le conseguenze delle attività umane non rientrano in categorie così pulite né sono così benevoli. Se le attività umane sembrano essere patologiche per l’ecosistema globale, dobbiamo trovare un modello più completo. Tra le possibilità vi sono le categorie patologiche di parassitismo, neoplasie o disturbi nella crescita, malattie infettive e processi infiammatori e disordini autoimmuni (Perez-Tamayo, 1961)
Nel rivedere queste alternative, il modello che include i “disturbi nella crescita” è immediatamente allettante poiché la crescita è la caratteristica più drammatica ed evidente della popolazione umana. La crescita della popolazione umana globale può essere caratterizzata in questo momento come non regolata e in rapida crescita. È più lento e moderatamente regolamentato in alcune località e persino in alcune regioni, ma è comunque inesorabile nella prospettiva globale. Nel primo Pleistocene, il tasso annuale di crescita della popolazione umana non superava probabilmente lo 0,001% comprese le fluttuazioni locali minori e temporanee (Hassan, 1981). La popolazione umana è raddoppiata ogni 100.000 anni circa. Nel Neolitico, la crescita della popolazione aumentò a un tasso dello 0,1% all’anno, raddoppiando ogni 700 anni (Saucier, 1972). Nell’anno 0 dell’era cristiana, la popolazione umana totale era di circa 250.000.000, ma è raddoppiata a 500 milioni entro il 1650. La peste nel 14° secolo ha rimosso un terzo della popolazione umana, provocando una temporanea riduzione del tasso di crescita, ma la popolazione poi raddoppiò nuovamente a 1,1 miliardi nel 1850, solo 200 anni dopo. La popolazione mondiale raggiunse i 2 miliardi di anni all’inizio degli anni ’30 e i 2,5 miliardi nel 1950. La popolazione umana ora raddoppia ogni 35-40 anni. Ogni anno aggiungiamo tra 90 e 100 milioni di persone alla popolazione mondiale. Raffigurare questa crescita estremamente rapida su una scala aritmetica che include il Pleistocene o anche le ultime migliaia di anni mostra una curva di crescita simile a quella osservata in altre popolazioni poco prima che crollino. Eppure gli umani hanno mostrato una straordinaria resistenza alle forze che causano il collasso di altre specie.
La crescita rapida e incontrollata è una caratteristica principale del tumore maligno. Le neoplasie maligne presentano quattro caratteristiche principali:
- Crescita rapida e incontrollata
- Invasione e distruzione di tessuti normali adiacenti
- De-differenziazione
- Metastasi a diversi siti (Anderson, 1961; Perez-Tamayo, 1961)
I tumori mostrano anche ciò che i biologi del cancro chiamano progressione, o un’evoluzione verso una maggiore malignità, con minore differenziazione delle cellule, crescita più rapida e cellule più metastaticamente aggressive (David Prescott, comunicazione personale, Ruddon, 1987).
CARATTERISTICHE MALIGNE DELLA SPECIE UMANA
Ora possiamo esaminare quelle caratteristiche della specie umana nel suo insieme che suggeriscono il confronto con un processo maligno. Il riconoscimento di una neoplasia dipende da un “gruppo di cambiamenti morfologici” e da comportamenti. Una neoplasia maligna è definita come “… una nuova crescita incontrollata di tessuto composto da componenti che hanno il potere di crescita e moltiplicazione relativamente liberi dalle solite restrizioni” (Anderson, 1961).
Crescita rapida e incontrollata
Non ci sono prove che la crescita complessiva della specie umana sia limitata in questo momento o lo sia stata per almeno 10.000 anni. Non vi è alcuna indicazione affidabile che la crescita della popolazione umana globale sarà limitata volontariamente nel prossimo futuro. Questo non vuol dire che gli umani non abbiano la capacità di regolare la crescita della popolazione. Al contrario, gli sforzi degli umani per controllare la loro fertilità sono diffusi, leggendari e pesantemente documentati (Devereux, 1955, Birdsell, 1957, 1968, Himes, 1970, Nag, 1962, Polgar, 1968, Dumond, 1975, Tinker et al., 1976; Hern, 1976). Gli uomini hanno regolato la loro fertilità nella preistoria e persino tra le popolazioni preindustriali contemporanee. Gran parte della nostra moderna crescita della popolazione non è il risultato di migliori cure mediche, ma delle interruzioni degli antichi controlli sulla fertilità e la natalità che hanno precedentemente mantenuto la crescita della popolazione a tassi più bassi. Ricerche recenti condotte su alcuni gruppi sudamericani indiani hanno dimostrato una fertilità estremamente elevata che può essere correlata alla rottura dei controlli tradizionali sulla fertilità e la natalità (Hern, 1988; Hem, 1990).
Nonostante gli sforzi per controllare la fertilità, la popolazione umana globale non è riuscita di recente come specie a farlo. Vi sono numerosi esempi di forze potenti che bloccano i tentativi di controllare la fertilità umana e persino di incoraggiare la fertilità. Questi vanno dai capi di stato e capi delle gerarchie religiose a gruppi di interesse privati. L’attuale presidente degli Stati Uniti, George Bush, ha giudicato che avrebbe potuto ottenere voti e migliorare le sue prospettive di essere eletto sostenendo l’imprigionamento di medici che praticano l’aborto (Boyd, 1988). Non ha ricevuto critiche efficaci e non ha pagato alcun prezzo politico per questa posizione. L’attuale Papa è famoso per andare in posti dove membri della popolazione locale in rapida crescita hanno spogliato la vegetazione originaria del paesaggio in uno sforzo disperato di nutrirsi e, al suo arrivo, li esorta a riprodursi il più possibile. Le forze pronataliste hanno, per la maggior parte, il controllo delle istituzioni umane. Una recente e drammatica inversione locale di questa tendenza fu il rovesciamento di Ceaucescu in Romania. Il primo atto del nuovo governo rivoluzionario fu quello di revocare il divieto di aborto e contraccezione da parte di Ceaucescu.
D’altra parte, il governo degli Stati Uniti, che ha una forte influenza sulle politiche e sui programmi della popolazione mondiale, ha annunciato alla Conferenza internazionale sulla popolazione di Città del Messico che la crescita della popolazione è “neutrale”, che le crisi di crescita demografica localizzate sono la prova di “troppo controllo e pianificazione governativa;” la crescita della popolazione dovrebbe essere rallentata da “meccanismi naturali” che deriveranno dallo “sviluppo” (Menken, 1986). L’attuale governo degli Stati Uniti ha ritirato il sostegno per il Fondo per la popolazione delle Nazioni Unite.
Una delle principali caratteristiche di una crescita cancerosa è che resiste alla regolamentazione. Una cellula cancerosa è una cellula che si riproduce senza limiti (Prescott & Flexer, 1986). I meccanismi regolatori che determinano il tasso di moltiplicazione cellulare e l’aumento della massa totale sono ostacolati al punto che la crescita non è controllata. Possono essere presenti meccanismi regolatori, ma non sono presenti in quantità sufficienti per essere efficaci, e alla fine vengono distrutti dal tessuto non regolato (Anderson, 1961).
Il fatto che alcuni umani limitino la loro fertilità in modo efficace, non si riproducano o sostengano la diffusa disponibilità di metodi di limitazione delle nascite non cambia il corollario fatto che la specie umana nel suo complesso non opera in questo momento alcun vincolo efficace riguardo alla crescita dei suoi numeri.
Invasione e distruzione dei tessuti normali adiacenti
Dall’inizio della storia registrata abbiamo visto esempi di distruzione umana degli ecosistemi globali che iniziano con la deforestazione del Mediterraneo (Darby, 1956, Angel, 1975, Jordan & Rowntree, 1986). Le foreste dei cedri del Libano furono distrutte per sempre per costruire le navi dei Fenici e il Tempio di Salomone. Esempi di distruzione dell’ecosistema locale sono innumerevoli.
I tumori si diffondono in due modi: vasta invasione e metastasi o colonizzazione a distanza. Le comunità umane, una volta stabilite, tendono a invadere e distruggere tutti gli ecosistemi adiacenti senza limiti. A causa dei limiti dell’antica tecnologia e delle effettive dimensioni della popolazione, e a causa delle tradizioni culturali che rispettavano gli ecosistemi naturali, la situazione non fu sempre così, ma sembra che ora stia accadendo con regolarità. Non esiste un ecosistema sul pianeta che non sia già distrutto, invaso o immediatamente minacciato in qualche modo dalla specie umana. In effetti, l’ecosistema globale è esso stesso sottoposto a considerevole stress dalle attività umane e si prevede che questo processo acceleri (Gentry, 1980: Sagan, 1981; Fyfe, 1981; Gomez-Pampa, et al, 1973; Moriarty, 1988; SCEP, 1970; Goudie, 1982; Hafele, 1980; Mooney, 1988; Repetto, 1989).
De-differenziazione
Per il patologo o l’oncologo, il termine “de-differenziazione” ha una varietà di significati, ma si riferisce principalmente al tipo di singola cellula. Una cellula normale ha un aspetto caratteristico che identifica il tipo di tessuto e persino l’organo da cui deriva. Il tessuto cardiaco ha l’aspetto del tessuto muscolare e può essere identificato come proveniente dal cuore. Le cellule colonnari che rivestono l’intestino o la cervice hanno apparenze tipiche. Le cellule tumorali tendono a perdere questo aspetto differenziato. Il patologo potrebbe essere in grado di determinare che una determinata cellula cancerosa proviene da tessuto adenomatoso ma potrebbe non essere in grado di dire se era originaria della mammella, dell’intestino o della ghiandola parotide o che un carcinoma a cellule squamose è originato dal polmone o da qualche altra parte.
A volte le cellule tumorali sembrano proprio cellule tumorali, ma la loro origine generale non è evidente. Meno una cellula sembra provenire da uno specifico tipo di tessuto, più è probabile che sia maligna; più è non identificabile, più è probabile che faccia parte di un tumore altamente maligno e aggressivo.
Una massa di tessuto canceroso può anche apparire indifferenziata nel senso che nessuna struttura specifica è visibile. Non c’è modo di determinare l’origine della cellula o dall’aspetto di singole cellule o dall’aspetto grossolano della lesione. A volte, per essere sicuri, particolari neoplasie hanno aspetti caratteristici, ma anche questi hanno perso le relazioni originali tra le cellule e le strutture che gli conferiscono un aspetto e una funzione normali.
Su base individuale, gli esseri umani sono notevolmente simili in anatomia, fisiologia e aspetto in tutto il mondo. Esistono differenze intraspecifiche minori che includono il colore della pelle, la statura, la configurazione facciale e specifici adattamenti locali al caldo o al freddo, o anche adattamenti a specifici problemi di malattia come il tratto falciforme che protegge dalla malaria da Plasmodium falciparum, ma le differenze rispetto ad altre specie adattate ai diversi ecosistemi sono assenti. Gli adattamenti culturali, invece degli adattamenti fisici, colmano il divario tra il !Kung Bushman nel deserto del Kalahari e suo fratello Nunamiut, l’eschimese cacciatore di foche del mare artico. Sono la stessa specie anche se non riescono a capire la lingua, il comportamento, la dieta o il modo di scelta del coniuge. Gli esseri umani sono “anaplastici” nel senso che non sono fisicamente molto differenziati tra loro, e l’aumento dei contatti tra tutti i gruppi accelera la perdita di differenze sia fisiche che culturali tra i singoli membri della specie.
In un senso più ampio, tuttavia, il fattore critico nell’anaplasia umana si trova nella capacità della cultura di eliminare l’importanza delle differenze negli adattamenti fisici o anche in adattamenti subculturali specializzati che erano precedentemente necessari per la sopravvivenza negli ecosistemi locali (Frisancho, 1981). Ora l’eschimese può usare una motoslitta invece di una slitta trainata da cani o di un kayak per raggiungere un sito di caccia dall’altra parte della baia. Gli adattamenti tecnologici occidentali permettono ai non-eschimesi di vivere nella regione del North Slope dell’Alaska con un comfort ragionevole in un clima e un ambiente sopravvissuto solo in precedenza da quelli immersi in una cultura altamente specializzata altamente adattata a quell’ambiente tanto rigido. Vado a vivere con i miei amici indiani Shipibo nella parte superiore dell’Amazzonia peruviana senza dover sapere nulla della caccia agli Arapaima gigas con un arpione o un cacciatore di pecari dal collare con arco e frecce. I miei adattamenti culturali, che porto con me, e la mia capacità di usare le competenze linguistiche e mediche per scambiare tipi di assistenza con lo Shipibo, mi permettono di vivere con loro confortevolmente per lunghi periodi di tempo. Mettimi nella giungla da solo senza attrezzatura e probabilmente non durerò a lungo. Viceversa, i miei amici Shipibo non sarebbero durati a lungo nell’inverno del Colorado senza gli adattamenti culturali locali che potevo fornire o insegnare loro a usare. Scambi negoziati di specifici adattamenti culturali, che riducono le differenze negli adattamenti locali, minimizzano le difficoltà umane nel sopravvivere in ambienti estremamente diversi.
A causa della loro mobilità e capacità di adattamento culturale, gli esseri umani possono sfruttare e sopravvivere in ecosistemi sparsi e diversi (Moran, 1982). A tale riguardo, gli esseri umani sono diversi dalla maggior parte delle altre specie di vertebrati che sono altamente adattati a nicchie ecologiche ristrette e specifiche. L’antropologo Joseph Birdsell (1968) osservò che l’uomo ha “abbandonato la densità per la plasticità” (pagina 248). L’antropologo William S. Laughlin (1968) ha osservato che, dopo aver trovato un cavallo, un uomo può farlo correre in due o tre giorni, quindi “decidere se mangiarlo, cavalcarlo, fargli tirare un carico, indossarlo o adorarlo “(p 313). Gli esseri umani sono ecologicamente più versatili di qualsiasi altra specie di vertebrati.
De-differenziazione a livello di comunità
Mentre si può argomentare che i singoli esseri umani sono membri “de-differenziati” di una singola specie ovunque presente, l’espressione della de-differenziazione nelle popolazioni umane può essere illustrata ad un altro livello di analisi, cioè il livello comunitario. Un visitatore dallo spazio potrebbe vedere non singoli esseri umani, ma lesioni sul paesaggio terrestre, alcuni con collegamenti interconnessi. Le lesioni ora brillano nell’oscurità (vedi la copertina di Scientific American di settembre 1989). Un osservatore spaziale osservando il pianeta negli ultimi tremila anni vedrebbe, in primo luogo, un’ampia varietà di queste lesioni con confini, schemi e strutture permanenti con forme uniche. L’architettura delle strutture permanenti sarebbe specifica, in molti casi, in posizioni distinte sul globo. La casa della comunità ovale di Yanomama sarebbe distintamente diversa da Kayapo o Waorani Ionghouse e questi ancora diversi dai modelli di architettura e residenza dei Dogon nell’Africa occidentale e dai trulli conici dell’Italia meridionale (Carver, 1979). Ora il viaggiatore internazionale trova file di case o appartamenti in apparenza identici a Zagabria, San Paolo, Levittown e Houston. Gli skyline di Chicago, San Paolo, Denver e Tokyo sono tra loro più simili che non differenti. Le apparizioni delle comunità umane su grande scala, in particolare nei centri urbani, stanno rapidamente scomparendo. Di fatto, a parte le differenze scalari, la morfologia degli schemi di insediamento delle grandi comunità umane non solo non si distinguono l’una dall’altra e sono indifferenziate nell’aspetto, ma diventano indistinguibili dalla morfologia delle lesioni maligne che si trovano in natura.
I tumori maligni sono caratterizzati da confini altamente irregolari e invasivi che sono indistinti. I tumori benigni tendono ad avere confini distinti e perfino incapsulati e non sono invasivi. Quando le immagini di lesioni maligne vengono confrontate con immagini di comunità umane, in particolare conglomerati urbani moderni e differenze scalari / tonali sono minimizzate o eliminate, come nelle Figure 11-15, le lesioni maligne e le comunità umane sono molto simili, anche indistinguibili, nelle loro apparenze. Questi due fenomeni molto diversi hanno la stessa morfologia. In azione ciascuno invade aggressivamente lo spazio adiacente e sostituisce il tessuto o l’ecosistema precedente con il proprio. Poiché le neoplasie maligne stabiliscono metastasi locali, anche le città sviluppano comunità satellite che alla fine diventano parte di un processo più ampio. Immagini aeree, immagini satellitari o immagini astratte di comunità umane e altre attività umane aziendali nel tempo in qualsiasi parte del pianeta illustrano questo fenomeno (Figura 16). È uno studio che dovrebbe prestarsi all’uso dei frattali come strumento di ricerca (La Breque, 1986/7).

FIGURE 11-15: Confronto delle somiglianze morfologiche tra lesioni metastatiche e communità umana.
a) adenocarcinoma polmonare; b) melanoma metastatico maligno; c) Baltimora; d) melanoma metastatico maligno; e) cluster a cinque città, North Carolina

FIGURA 16. Un confronto tra il mappatore tematico Landsat 1,55-1,75 μm di dati del 1984 e 1988 aumenta la deforestazione tropicale nel bacino amazzonico, nel Mato Grosso, in Brasile. Le aree disboscate appaiono più chiare, mentre le aree indisturbate della foresta tropicale appaiono più scure. (Fonte: Instituto Nacional de Pesquisas Espaciais)
Odum (1989) nota che le città sono ecosistemi altamente eterotrofi e parassiti poiché consumano molta più energia di quanta ne producano. Mentre le megalopoli della fine del XX e dell’inizio del XXI secolo si sviluppano, con numerosi centri urbani contenenti decine di milioni di abitanti, le grandi città del mondo si concentrano in sistemi eterotrofi che attingono energia da parti distanti del pianeta e creano un aumento quantico nell’entropia.
Metastasi
La metastasi, o la disseminazione in siti distanti, è una caratteristica primaria delle neoplasie maligne. In questo modo si formano neoplasie maligne in diversi sistemi di organi lontani dal tessuto o organo d’origine. È il processo attraverso il quale il cancro si diffonde più rapidamente e letalmente per uccidere l’organismo ospitante.
Il corollario delle metastasi nella popolazione umana è la creazione di nuove colonie umane in siti lontani dalle origini dei loro membri. Le capacità migratorie degli esseri umani sono sbalorditive. Ancora più sorprendente è la capacità degli esseri umani di trovare un nuovo ecosistema, identificare le componenti benefiche e dannose del sistema e adattare la loro cultura per soddisfare i bisogni di sopravvivenza nel nuovo sistema. La foresta pluviale amazzonica, ad esempio, è un ambiente estremamente complesso, ma gli amazzoni autoctoni hanno vissuto con successo lì per almeno 10.000 anni (Lathrap, 1970; Roosevelt, 1980; Posey, 1983). Stavano andando bene fino a quando non furono contattati dagli esploratori europei (Denevan, 1976; Myers, 1989; Hecht & Cockburn, 1989; Hern, 1990). Crosby (1988) ha documentato l’espansione della cultura europea attraverso le colonie e la distruzione che ha portato agli ecosistemi del Nuovo Mondo. Insieme a questi contatti arrivarono devastanti epidemie e perdite fino al 95% delle popolazioni indigene (Dobyns, 1983) seguite da una rapida espansione delle popolazioni coloniali. Anche Malthus (1798) ha commentato la rapida crescita delle popolazioni nelle colonie americane. Le conseguenze dell’espansione coloniale sono simili in molti modi al cancro metastatico altamente aggressivo.
Criteri per la diagnosi
Delle quattro caratteristiche dei tumori, i patologi generalmente richiedono che un tumore ne mostri due per essere classificato come maligno. Inoltre, il comportamento di un tumore è più importante della sua istogenesi (Perez-Tamayo, 1961).
La mia ipotesi è che la popolazione umana abbia tutte e quattro le caratteristiche di un processo maligno e che il suo comportamento in almeno tre categorie sia chiaramente maligno (Tabella 1).
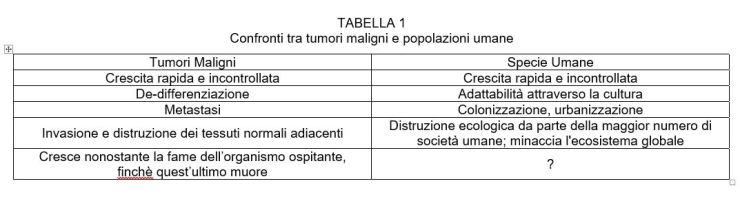
Descrizione e diagnosi
La specie umana è una specie rapace, predatrice, omniecofagica, impegnata in un modello globale di conversione di tutta la materia disponibile vegetale, animale, organica e inorganica nella biomassa umana o in complementi funzionali alla biomassa umana. Questo è un processo epiecopatologico ecocida, sia nell’immediato che in futuro.
A tale riguardo, la specie umana è un esempio di ecotumore maligno, una proliferazione incontrollata di una singola specie che minaccia l’esistenza di altre specie nei loro habitat. Un gigantesco stormo di storni o merli può essere un esempio di un ecotumore benigno che è odioso per gli umani, ma è improbabile che lasci alcun danno duraturo. Un esempio di un altro ecotumore maligno, tuttavia, è la corona di specie di stelle marine spinose che sta distruggendo la Grande Barriera Corallina nel Pacifico meridionale (Yonge, 1963). Ancora un altro è l’anguilla lampreda, che ha ottenuto l’accesso ai Grandi Laghi attraverso il canale St. Lawrence e ha distrutto il pesce nativo dei Grandi Laghi (Benard, 1989). Entrambi sono il risultato di meccanismi di regolazione disturbati o di interruzioni antropogeniche all’interno dell’ecosistema (Moriarty, 1988). Nel caso della lampreda, gli umani hanno iniziato a invertire il danno ecologico mediante l’applicazione di una tecnologia sofisticata.
Discussione
Una forma altamente maligna di cancro resiste a tutti gli sforzi per una regolamentazione efficace fino alla morte dell’organismo ospitante. I tumori cancerosi continuano a crescere anche di fronte alla fame dell’ospite (Ruddon, 1987).
Nel caso della specie umana, questa ipotesi prevede che l’ecosistema globale, il substrato di supporto, debba essere distrutto prima che la neoplasia venga arrestata, se, in effetti, si arresta. Una proprietà peculiare della specie umana è che è geniale nei suoi adattamenti culturali a fronte di difficili problemi di sopravvivenza. Ora si sta persino spostando oltre l’atmosfera terrestre nello spazio, un ambiente che è ostile a tutte le forme di vita. Sotto questo aspetto, possiamo vedere il primo sbarco sulla luna come metastasi anticipatorie.
Gli esseri umani sono sopravvissuti e hanno prevalso non solo imparando a sfruttare tutti gli ecosistemi e le loro componenti viventi, ma imparando a sfruttare altre risorse organiche e inorganiche come petrolio, gas naturale e minerali. La specie umana è esperta nel trovare modi per convertire le sostanze più improbabili sia per la produzione di cibo sia per l’adattamento degli ambienti necessari alla vita dell’uomo. C’è da chiedersi se l’eliminazione dell’intero ambiente naturale impedirebbe la sopravvivenza della specie umana, tenuto conto della capacità della specie di adattarsi o di convertire le risorse al suo uso.
È possibile, naturalmente, che la specie umana riconosca ciò che sta facendo al pianeta in tempo per invertire la tendenza e ripristinare l’ecosistema naturale alle condizioni precedenti dell’intervento umano. È anche possibile che la specie umana riesca a continuare a crescere convertendo tutti i materiali del pianeta ad uso umano. Un problema importante, avuto presente che la specie umana si è evoluta in ecosistemi complessi. Siamo abbastanza intelligenti da gestire un ecosistema abbastanza complesso da sostenerci? “Cercare di rendere la natura stabile non ti porta da nessuna parte” (Schaffer, 1990). Per parlare della sola atmosfera, stiamo conducendo un esperimento incontrollato di quello che potrebbe rivelarsi il più grande garage chiuso dell’universo conosciuto.
Una delle principali tendenze della specie umana è quella di semplificare gli ecosistemi ovunque. Gli ecosistemi semplificati sono intrinsecamente instabili e tendono a collassare. Abbiamo notevolmente semplificato le nostre relazioni preda-predatore; questo aumenta la probabilità di sperimentare oscillazioni acute e incontrollabili sia nei nostri sistemi di supporto vitale che nei nostri livelli di popolazione. La cultura umana ha rimosso i vincoli che determinano oscillazioni smorzate caratteristiche della maggior parte delle specie. Non abbiamo ancora conosciuto o preso coscienza di alcun anello di feedback negativo che metta seriamente in pericolo la nostra sopravvivenza come specie anche se ci appare davanti. Il risultato è una crescita non regolata e un’instabilità potenzialmente letale della popolazione che degenera in caos figurativo, letterale e matematico.
La teoria del caos prevede che le oscillazioni non campionate procedano all’estinzione in funzione di crescenti tassi di crescita (maggio 1974, maggio 1976, maggio 1976, May & Oster, 1976, Southwood, 1976, Schaffer e Kot, 1985). Piccoli cambiamenti nelle condizioni iniziali, specialmente nei tassi di crescita, possono provocare oscillazioni ampie, deterministiche ma imprevedibili nei cicli di popolazione futuri. La nostra esperienza con altre specie biologiche e la nostra storia recente dovrebbe dirci che siamo pericolosamente vicini al caos irrecuperabile se non abbiamo già stabilito da molto tempo il modello che porterà a quel risultato. È possibile che l’aumento della crescita della popolazione umana dallo 0,001% all’anno allo 0,1% all’anno alla fine del Paleolitico ci ha portato ad entrare in un regime caotico deterministico che si estende per migliaia di anni e che non possiamo prevedere? È probabile che eviteremo completamente le restrizioni ecologiche sperimentate da altre specie (Woodwell, 1985)? È probabile che, per gli umani, non ci sia un limite alla “capacità di carico” dell’ecosistema terrestre? Se c’è un limite, quanto tempo ci vorrà per raggiungerlo (von Foerster et al, 1960)? Cosa accadrà quando lo faremo?
Lo scopo di ogni ipotesi è di spiegare la realtà e prevedere gli eventi. L’ipotesi che la specie umana sia una neoplasia maligna globale, un processo ecopatologico, fornisce una spiegazione unificante di un’ampia varietà di eventi e fenomeni tra cui una rapida crescita della popolazione umana, una diffusa distruzione umana di ambienti locali, regionali e continentali, nonché dell’ecosistema globale, rapida urbanizzazione, cambiamenti atmosferici globali, estese estinzioni di specie di natura antropogenica, resistenza alla regolazione della popolazione e numerose altre osservazioni. È un fenomeno potenzialmente reversibile e quindi un’ipotesi confutabile. La specie umana è in grado di regolare la sua fertilità e la crescita della popolazione, è in grado di ripristinare ambienti e di salvare altre specie dall’estinzione ed è in grado di vivere in armonia con il resto dell’ecosistema.
Questa ipotesi prevede che, mentre la specie umana è capace di tutte queste attività non cancerose e anche occasionalmente le visualizza, continuerà a comportarsi complessivamente come un cancro sul pianeta.
Forse il modello del caos è più esplicativo del modello di una neoplasia globale. Forse è una combinazione di questi, o, come affermano Brooks and Wiley (1986), la specie è una manifestazione della tendenza evolutiva nella direzione di una maggiore entropia. Polgar (1961) affermò che l’evoluzione ritarda l’entropia, mentre i sistemi umani epici aumentano la complessità delle strutture che modificano sempre più l’ambiente naturale: “… gran parte della superficie terrestre fa parte di un singolo sistema ecologico unificato dalla specie umana e gli organismi ad esso associati ” (p 105). Praticamente tutti gli esseri umani aumentano l’entropia (Polgar, 1961; Goldman, 1970). Le guerre, in particolare la guerra del Vietnam, hanno provocato grandi aumenti nell’entropia (Orians & Pfeiffer, 1970). L’inefficienza biologica e gli adattamenti culturali degli esseri umani fanno affondare la specie nell’entropia, con la comunità urbana come una sorta di punto di concentrazione nodale di entropia. Forse il pianeta ha, nella specie umana, un caso di entropia maligna, con il riscaldamento globale come una delle manifestazioni, un po’ come la malaria o il paziente colpito da un colpo di calore con un caso di “iperpiressia maligna” (Bruce-Chwatt, 1971).
La cultura umana è al tempo stesso una delle principali fonti globali di disturbo normativo dell’ecosistema e l’unica speranza, a meno dell’estinzione umana, per invertire il processo e ripristinare l’ecosistema.
Una nuova specie
Per descrivere il fenomeno di questa specie maligna, omniecofagica, propongo che il nuovo nome scientifico della specie umana sia l’homo ecophagus (homo = man (L.); oikos = casa, anche la radice tradizionale per “ecosistema” (Gr.); phagos = ghiottone (Gr.)) – “l’uomo che divora l’ecosistema”.
Propongo che il termine generale che dovrebbe essere usato per l’homo ecophagus sia “Protoneontos”. Potremmo aspirare a un nuovo stato di coscienza globale relativamente alla necessità di preservare il nostro ecosistema comune; sarebbe un allontanamento importante dal comportamento umano tradizionale, un “nuovo essere” – Neontos (neos = nuovo, fresco, anche strano, inaspettato (Liddell & Scott, 1958) (Gr.) e ontos = essere (Gr.). Protoneontos, il primo dei nuovi esseri e antecedente a Neontos, sta distruggendo l’ecosistema planetario. Il precursore di Protoneontos, Paleoneontos, conosciuto anche come homo sapiens, “uomo saggio”, non lo fece.
La diagnosi finale
La nuova specie umana, l’homo ecophagus, è una specie onnipresente, predatrice, omniecofagica, un processo epiecopatologico maligno impegnato nella conversione di tutto il materiale planetario in biomassa umana o nel suo sistema di supporto con squilibrio terminale dell’ecosistema globale.
Conclusione
L’idea che la popolazione umana sia un cancro planetario è una conclusione profondamente inquietante, ma le osservazioni della comunità scientifica degli ultimi 20 anni hanno fornito un sostegno massiccio a questa ipotesi e poco, se non altro, per confutarla. È estremamente raro che qualsiasi cancro si arresti volontariamente e spontaneamente. Se la specie umana può smettere di essere un cancro, sarà una di quelle eccezionali rarità, o la mia ipotesi è sbagliata. Accetterò volentieri entrambi i risultati.
______________________________________________________________________________________
Articolo originale pubblicato su “Population and Environment: A Journal of Interdisciplinary Studies, volume 12, numero 1, autunno 1990

2 pensieri riguardo “Perchè siamo così tanti ?”